Amino acid metabolism and signaling in plants
Metabolic pathways
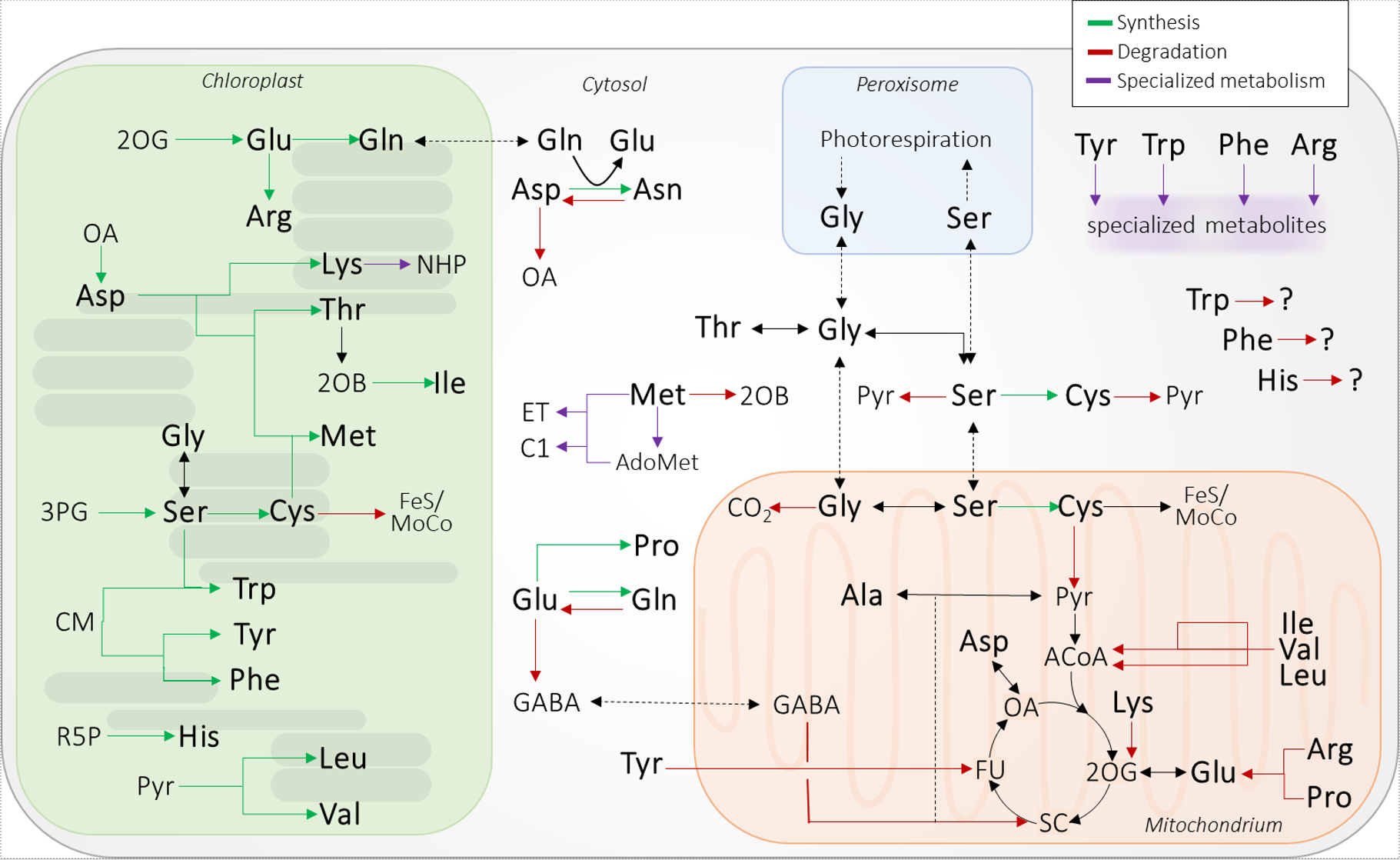
Plants are able to synthesize all 20 amino acids required as building blocks for proteins and in addition they produce a diverse set of non-protein amino acids and secondary metabolites. Amino acid synthesis pathways are mainly localized in the chloroplasts and require precursors as well as energy provided by photosynthesis (Fig. 1). The individual reaction steps involved in amino acid synthesis and their regulation have been extensively studied. In contrast, the catabolic pathways in plants are still not completely clear (Hildebrandt et al. 2015). Our group identified the mitochondrial pathway for cysteine degradation (Krüßel et al. 2014, Höfler et al. 2016). Cysteine metabolism in plants is exceptional in that it is distributed across different compartments. Several isoforms of cysteine synthesizing enzymes are present in chloroplasts, mitochondria and the cytosol and there are also diverse catabolic pathways. Interstingly, this compartmentalization of cysteine metabolism seems to be critical for its signaling functions, which are just beginning to emerge (Heinemann and Hildebrandt 2021). Recently there has been some major progress achieved by several independent groups in elucidating missing steps in the degradation pathways for the branched-chain amino acids and lysine (Heinemann and Hildebrandt 2021). However, several amino acid catabolic pathways in plants remain unresolved.
Further reading:
Heinemann B and Hildebrandt TM (2021) The role of amino acid metabolism in signaling and metabolic adaptation to stress-induced energy deficiency in plants. J Exp Bot 72, 4634-4645. doi: 10.1093/jxb/erab182
Hildebrandt TM, Nunes Nesi A, Araújo WL, Braun HP (2015) Amino Acid Catabolism in Plants. Mol Plant 8: 1563–1579. doi: 10.1016/j.molp.2015.09.005
Höfler S, Lorenz C, Busch T, Brinkkötter M, Tohge T, Fernie A, Braun H.P., Hildebrandt TM (2016) Dealing with the sulfur part of cysteine: four enzymatic steps degrade l-cysteine to pyruvate and thiosulfate in Arabidopsis mitochondria. Physiol Plant 157, 532-366. doi: 10.1111/ppl.12454
Krüßel L, Junemann J, Wirtz M, Birke H, Thornton JD, Browning LW, Poschet G, Hell R, Balk J, Braun HP, Hildebrandt TM (2014) The mitochondrial sulfur dioxygenase ETHE1 is required for amino acid catabolism during carbohydrate starvation and embryo development in Arabidopsis thaliana. Plant Physiol 165, 92-104. doi: 10.1104/pp.114.239764
Metabolic plasticity
Plants are confronted with a broad spectrum of unfavorable environmental conditions ranging from abiotic stresses such as drought, flooding, high salinity, or extreme temperatures to attacks by various pathogens. Being sessile, plants had to develop a sophisticated metabolic defense system to differentiate between the various threats, correctly assess them, and adapt their defense strategy accordingly. Amino acids are required as precursors for the synthesis of stress related proteins, secondary metabolites and signaling molecules. In addition, the oxidation of amino acids as alternative respiratory substrates provides ATP during energy deprivation, which may occur in the context of environmental stress but also during development. Nitrogen and its allocation pattern might be a pivotal component of trade-offs between growth and stress tolerance in plants. Proteins are by far the largest nitrogen sink in plants, and protein metabolism including amino acid synthesis as well as protein turnover accounts for a significant proportion of the energy required for plant growth. Our group has studied the intersection of plant amino acid metabolism and abiotic stress response on a qualitative and quantitative level (Heinemann et al. 2021, Batista-Silva et al. 2019, Hildebrandt 2018). We are currently using germination, hypoxia, drought, and energy deficiency induced by extended darkness as model conditions to investigate amino acid metabolic plasticity in Arabidopsis thaliana and Lupinus albus (Angermann et al. 2024).
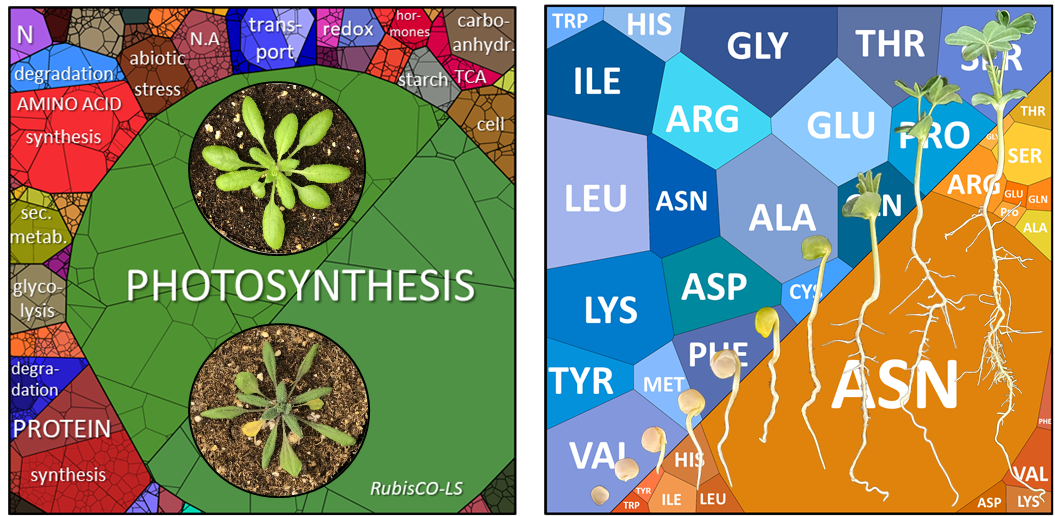
The Voronoi diagrams (Fig. 2) were generated here.
Further reading:
Angermann C, Heinemann B, Hansen J, Töpfer N, Braun HP, Hildebrandt TM (2024) Proteome reorganization and amino acid metabolism during germination and seedling establishment in Lupinus albus. Journal of Experimental Botany. doi: 10.1093/jxb/erae197
Batista-Silva W, Heinemann B, Rugen N, Nunes-Nesi A, Araújo WL, Braun HP, Hildebrandt TM (2019) The role of amino acid metabolism during abiotic stress release. Plant Cell Environ 42, 1630-1644. doi: 10.1111/pce.13518
Heinemann B, Künzler P, Eubel H, Braun HP, Hildebrandt TM (2021) Estimating the number of protein molecules in a plant cell: protein and amino acid homeostasis during drought. Plant Physiol 185, 385–404. doi: 10.1093/plphys/kiaa050
Hildebrandt TM (2018) Synthesis versus degradation: directions of amino acid metabolism during Arabidopsis abiotic stress response. Plant Mol Biol 98, 121-135. doi: 10.1007/s11103-018-0767-0
Metabolic interactions
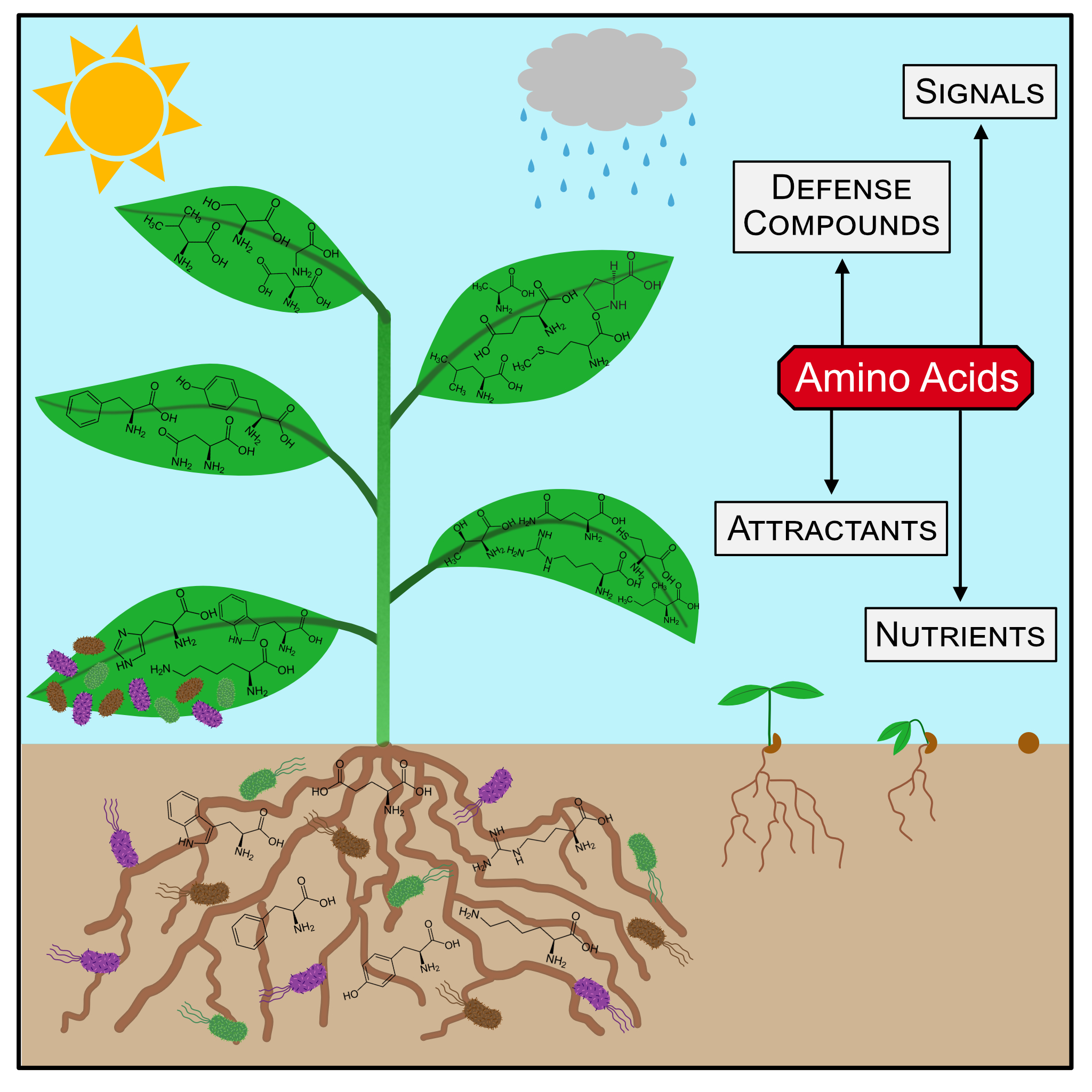
Plants constantly get into contact with a diverse mix of pathogenic and beneficial microbes. The ability to distinguish between them and respond appropriately is essential for plant health. We are interested in understanding the role of amino acid sensing, signaling, transport, and metabolism during plant-microbe interactions. Biochemical pathways converting individual amino acids into active compounds were recently elucidated and comprehensive large-scale approaches brought amino acid sensors and transporters into focus (Moormann et al. 2022). These findings show that plant primary amino acid metabolism is closely interwoven with stress signaling and defense responses at various levels. The individual biochemical mechanisms and the interconnections between the different processes are just beginning to emerge and might serve as a foundation for new plant protection strategies.
Further reading:
Moormann J, Heinemann B, Hildebrandt TM (2022) News about amino acid metabolism in plant-microbe interactions. Trends in biochemical sciences 47 (10):839–850. doi: 10.1016/j.tibs.2022.07.001
Amino acid sensing and signaling
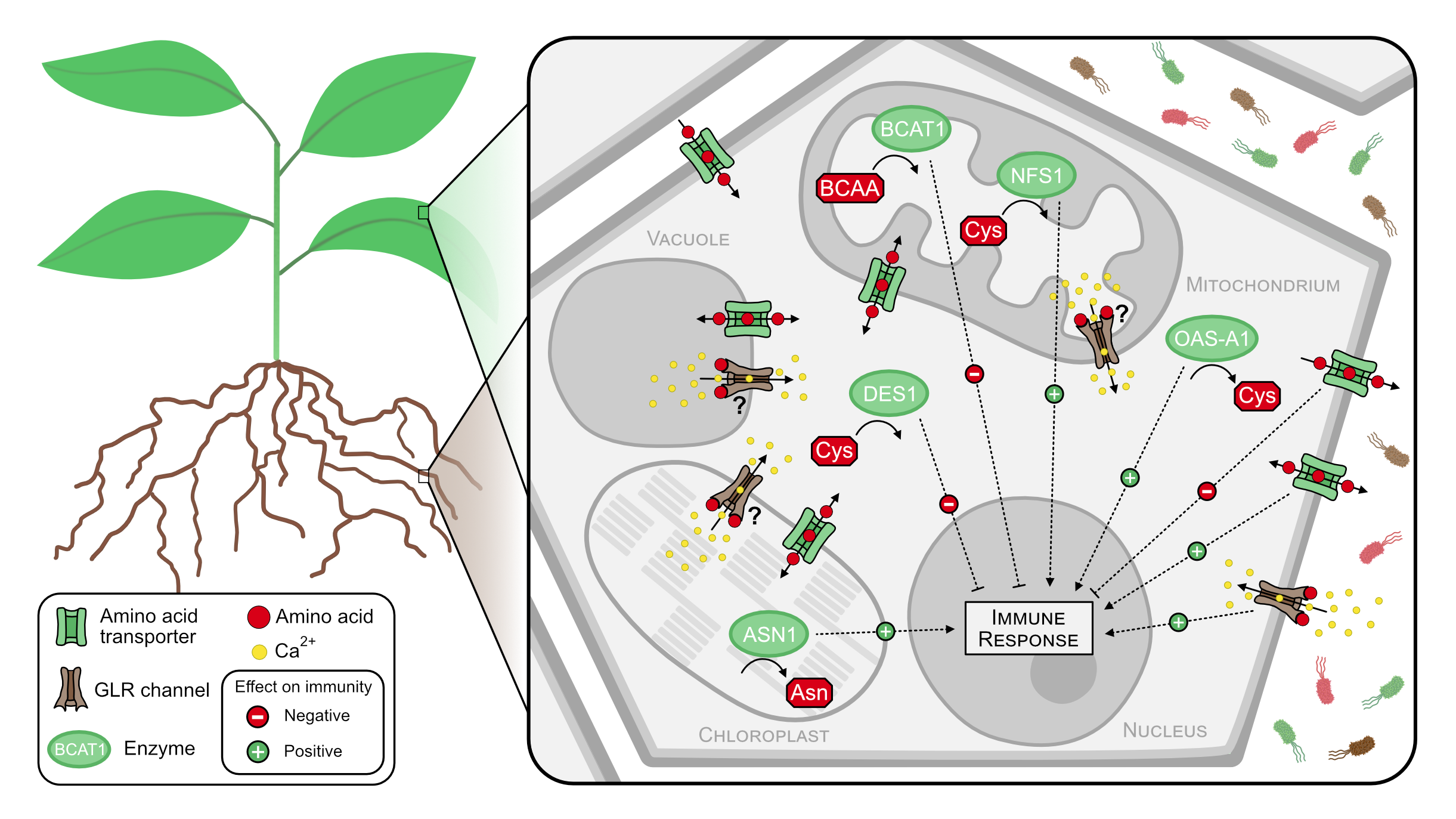
Potential mechanisms of amino acid signaling and perception include post-translational protein modifications (PTMs). PTMs can activate or repress a protein function and also affect protein stability. Reversible protein persulfidation as a regulation mechanism requires the attachment and removal of a persulfide (-SSH) group to a cysteine residue. Specific functions of persulfidation in plants have already been shown during drought response and autophagy (Heinemann and Hildebrandt 2021) but the mechanism of this regulation is still largely unknown. We are studying persulfidation as a post-translational protein modification and its role during plant microbe interactions.
Further reading:
Heinemann B and Hildebrandt TM (2021) The role of amino acid metabolism in signaling and metabolic adaptation to stress-induced energy deficiency in plants. J Exp Bot 72, 4634-4645. doi: 10.1093/jxb/erab182
Moormann J, Heinemann B, Hildebrandt TM (2022) News about amino acid metabolism in plant-microbe interactions. Trends in biochemical sciences 47 (10):839–850. doi: 10.1016/j.tibs.2022.07.001